
Das Jahr 1958 war eines der bedeutendsten Jahre für die Geschichte der Molekularbiologie. In den Jahren davor beschäftigten sich hauptsächlich Chemiker und Mediziner mit den bedeutenden Molekülen des Lebens. Friedrich Miescher, Albrecht Kossel und Erwin Chargaff lieferten erste wichtige Erkenntnisse über das Vorhandensein und die chemische Zusammensetzung der DNA. Oswald T. Avery erkannte die Bedeutung der DNA als das Molekül, welches unsere Erbinformation speichert und an die nächste Generation weitergibt. Und Anfang des Jahres 1953 waren es schließlich James Watson, Francic Crick, Maurice Wilkins und Rosalind Franklin, welche die Struktur dieser entschlüsselten. Es gab im Anschluss drei Vorstellungen, wie sich die DNA verdoppeln könnte: die konservative, die semikonservative und die disperse Replikation.
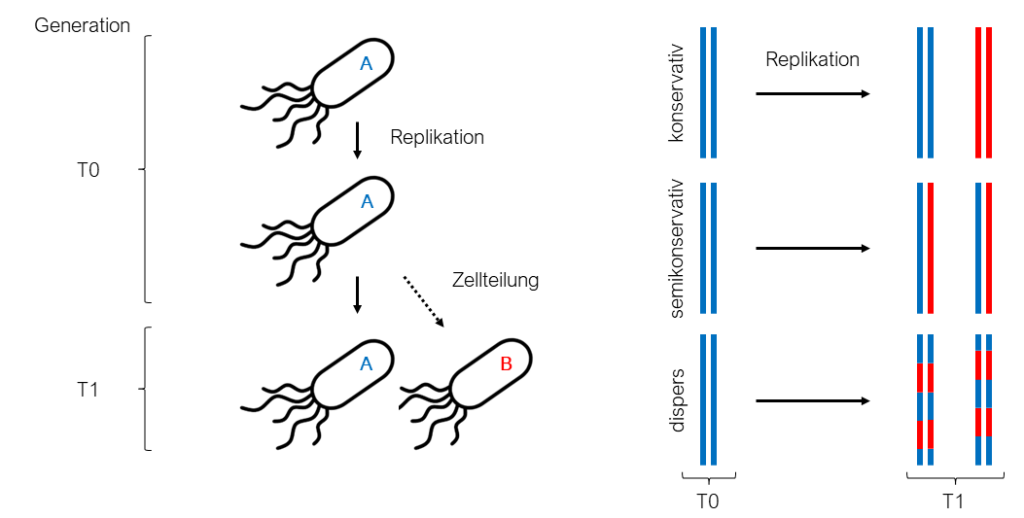
1. Konservative Replikation: Die T1-DNA bleibt unverändert und eine zusätzliche Kopie dieser wird synthetisiert.
2. Semikonservative Replikation: Die Wasserstoff-Brücken-Bindungen, welche die beiden Einzelstränge der Mutter-DNA (T1-DNA) zusammenhalten, werden aufgetrennt. Unter Beachtung der Chargaff´schen Regeln synthetisiert die Zelle für jeden der beiden vorliegenden Einzelstränge das komplementäre Gegenstück.
3. Disperse Replikation: Die T1-DNA teilt sich in wie bei der semikonservativen Replikation in ihre zwei Stränge und zusätzlich in kleinere Teile. Komplementär zu diesen wird von der Zelle das Gegenstück synthetisiert. In der T2-DNA besteht so aus einer Mischung aus T1-DNA und der neusynthetisierten DNA.
Um nun herauszufinden, welche dieser Theorien der Wirklichkeit entspricht, haben sich Matthew Messelson und Franklin Stahl, einen Versuch überlegt, welcher auf dem Einbau verschiedener Stickstoffisotope in die DNA bei der Replikation basiert. Kurz zu den Grundlagen: Alle bekannten Elemente sind in einer Tabelle, dem Periodensystem, anhand ihrer Ordnungszahl angeordnet. Die Ordnungszahl gibt an, wie viele Protonen sich in dem Atomkern jedes Elements aufhalten. Neben den Protonen bestehen Atomkerne zudem noch aus Neutronen. Während sich die Anzahl an Protonen für ein Element nicht ändern kann (ansonsten würde es sich um ein anderes Element handeln) kommen in der Natur verschiedenste Formen desselben Elements mit jedoch unterschiedlicher Neutronenzahl vor. Atome mit einer anderen Anzahl an Neutronen wie üblich, werden Isotope genannt. Stichstoff (N) ist eines der am häufigsten verbreiteten Elemente der Erde und kommt so u. a. auch als atomarer Bestandteil der DNA vor. Stickstoff hat die Ordnungszahl 7 und kommt zu 99% mit 7 Neutronen im Kern vor. Daher wird es auch als 14N bezeichnet (7 Protonen + 7 Neutronen => eine Massenzahl von 14). Zu den übrigen 1% gehört u. a. das Stickstoff Isotop 15N, welches ein Neutron mehr enthält als 14N (7 Protonen + 8 Neutronen => Massenzahl von 15).
Messelson und Stahl begannen damit, Escherichia coli (E.-coli-Bakterien) in einem Medium anzuziehen, welches das Stickstoff-Isotop 15N enthält. Die Bakterien, sind dementsprechend gezwungen, das vorhandene 15 N in ihrer DNA einzubauen. Ein E.-coli-Bakterium teilt sich unter guten Bedingungen alle 20 Minuten, aus einem Bakterium werden so zwei. Somit muss sich auch die DNA alle 20 Minuten verdoppeln, damit sich sowohl in dem Mutter-Bakterium (T1) als auch in dem neu entstandenen Bakterium (T2) eine Kopie des gesamten Genoms vorliegt (In der Abbildung ist das Mutter-Bakterium mit „A“ und das neu entstandene Bakterium mit „B“ beziffert). Messelson und Stahl nahmen nun E.-coli-Bakterien, deren DNA das Stickstoff-Isotop 15N enthält, und gaben diese in ein Medium, welches 14N anstelle von 15N beinhaltet. Die Folge dessen ist, dass sich die Bakterien bei weiterer Teilung, und der damit verbundenen Replikation der DNA, von dem 14N bedienen müssen und es in die DNA einbauen. Geht man von einer konservativen Replikation der DNA aus, hätte man nach 20 Minuten DNA in gleichen Mengen vorliegen, die entweder 15N oder 14N enthält. Bei der Annahme einer semikonservativen oder dispersen Replikation würde zum selben Zeitpunkt DNA vorliegen, welche in gleichen Mengen sowohl 15N als auch 14 N enthält. Um nun herauszufinden, welche der beiden Möglichkeiten vorliegt, wurde die DNA der Bakterien isoliert und mit Hilfe einer Dichte-Gradienten-Zentrifugation analysiert. Bei dieser Methode werden Substanzen in einem Gemisch nach ihrer Masse und Größe aufgetrennt. Hierbei wird die aufzutrennende Substanz auf einen Gradienten gegeben. Mit einem Gradienten ist hier ein Lösemittel gemeint, welches in einem Gefäß eine von unten nach oben abnehmende Konzentration aufweist. Wird dieses Gefäß mit dem Gradienten und der aufzutrennenden Substanz anschließend lang genug zentrifugiert, migriert die Substanz durch diesen Gradienten von oben nach unten und die sich darin befindenden Moleküle bleiben an der für ihre Größe und Masse spezifische Stelle des Gradienten stehen.
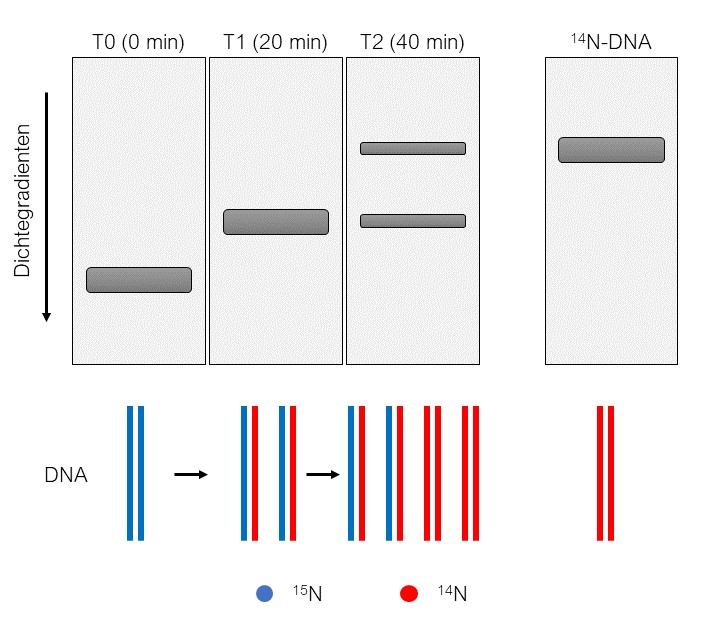
Die 15N-DNA ist aufgrund des zusätzlichen Neutrons in jedem Stickstoff-Atom schwerer als die 14N-DNA und migriert somit weiter in Richtung Boden des genutzten Dichte-Gradienten. So kann man einfach unterscheiden, ob man in einem Gemisch aus DNA 15N- und/oder 14N-DNA vorliegen hat. Zum Zeitpunkt 0 (T1 Generation) zeigte die Dichte-Gradienten-Zentrifugation lediglich eine Bande (Mit Bande ist das Signal gemeint, welches aufgrund des Vorhandenseins von DNA an dieser Stelle entsteht). DNA wiederum, die aus den Bakterien der T2-Generatuin isoliert wurde, welche für 20 Minuten in dem 14N-Medium aufgewachsen sind, zeigte nach der Zentrifugation ebenfalls eine Bande, die jedoch höher erscheint als die vorherige. Was bedeutet das? Ein höher verlaufendes Signal bedeutet, dass die DNA nicht ausschließlich das schwerere 15N eingebaut hat, sondern nun auch das leichtere 14N. Dass es sich dabei zusätzlich nur um nur eine Bande und nicht um zwei handelt, bedeutet, dass nur eine Form von DNA vorliegt, und zwar eine, welche sowohl 15N als auch 14N enthält! Damit konnte man zu diesem Zeitpunkt bereits die Theorie der konservativen Replikation ausschließen. Nach dieser hätte man nämlich zu diesem Zeitpunkt das Auftauchen zweier Signale erwartet: ein Signal auf gleicher Höhe wie bei der T1-Generation (DNA enthält nur 15N) und ein noch höherverlaufendes Signal (DNA enthält nur 14N). Um die Theorie zu bestätigen, wurde dieser Vorgang nun wiederholt: Man isolierte DNA aus Bakterien, welche für 40 Minuten (aufgrund der zwei Generationen) im 14N-Medium wuchsen und nutze erneut die Dichte-Gradienten-Zentrifugation, um die DNA aufzutrennen. Wie zu erwarten war, erschienen diesmal zwei Banden mit gleicher Intensität (die Intensität der Bande ist proportional zu der Menge an DNA): ein Signal auf der Höhe der 14N-Referenz und ein weiteres auf derselben Höhe wie zuvor. Dieses Ergebnis unterstützt die Annahme, dass es sich bei der Replikation der DNA nicht um eine konservative Replikation handelt. Es ist jedoch nicht ausreichend dafür, eine Aussage zu treffen, ob die DNA nun semikonservativ oder dispers repliziert wird.
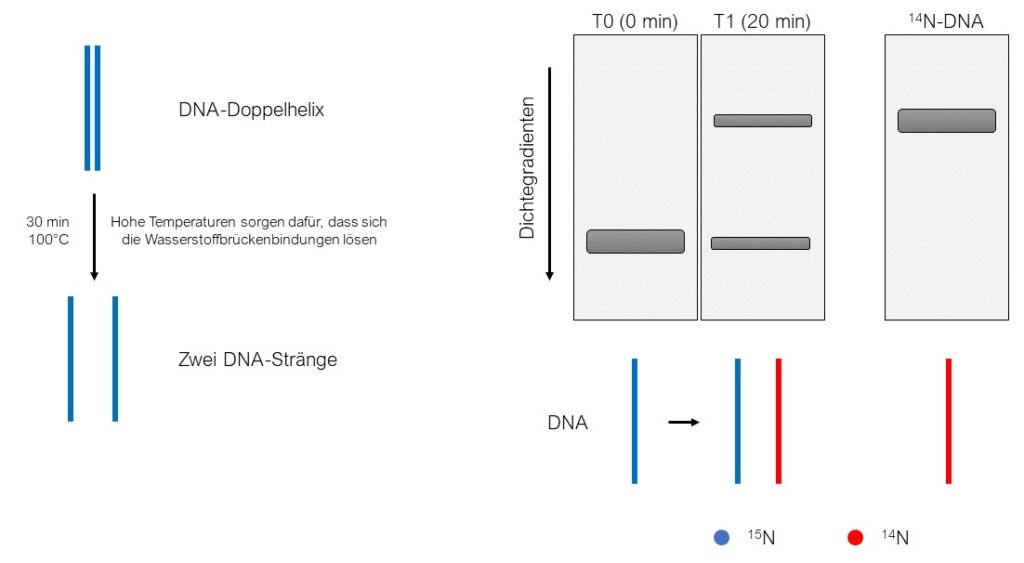
Um diese Frage zu beantworten, entnahmen die Wissenschaftler erneut DNA aus Bakterien der T1-Generation. Zur Erinnerung: In dieser befanden sich 15N und 14N in gleichen Mengen. Messelson und Stahl entschieden sich, die isolierte T1-DNA für einige Minuten der Temperatur von 100°C auszusetzen. Dies hatte zur Folge, dass die Wasserstoff-Brücken-Bindungen, welche die beiden Einzelstränge der DNA zu einer Doppelhelix zusammenhalten, aufgebrochen wurden. Mit der einzelsträngigen DNA wurde erneut eine Dichte-Gradienten-Zentrifugation durchgeführt. Es erschienen zwei Signale, die denen von erhitzter 15N- und 14N-DNA entsprachen! Daraus konnte geschlussfolgert werden, dass sich die DNA semikonservativ und nicht dispers repliziert. Würde sich die DNA dispers replizieren, hätte man zu diesem Punkt nach der Dichte-Gradienten-Zentrifugation ein Signal erwartet, welches mittig zwischen der einzelsträngigen 15N- und der einzelsträngigen 14N- DNA verläuft.
Zusammengefasst konnten Matthew Messelson und Franklin Stahl mit einem simplen Versuchssaufbau einen zu dem Zeitpunkt unerforschten Bereich der Molekularbiologie aufdecken und erste Erkenntnisse zum Vorgang der Replikation der DNA liefern. Ihre Arbeit war von großer Bedeutung, sodass viele WissenschaftlerInnen in den darauffolgenden Jahren weiter an dem Thema der DNA-Replikation forschten, um die genauen und grundlegenden molekularen Mechanismen dieses Vorganges aufzuklären. Um ein paar bedeutende Namen zu erwähnen: Arthur Kornberg und Severo Ochoa gelang es ein Enzym, die DNA-Polymerase I, zu isolieren, welche die Reaktion der Replikation katalysiert. Hierfür bekamen beide im Jahre 1959 den Nobelpreis für Medizin verliehen. Weitere wichtige Proteine, die in dem Prozess der Replikation involviert sind wurden entdeckt von: Hartmut Hoffmann-Berling (Helikase), Bernard Weiss und Carles Richardson (Ligase) und James Wang (Topoisomerase). Die Japanische Molekularbiologin Tsuneko Okazaki entdeckte mit ihrem Mann, Reiji Okazaki, die „Okazaki-Fragmente“. Kary Mullis publizierte zusammen mit Michael Smith im Jahre 1983 die Idee der Polymerase-Kettenreaktion (PCR) welche auf die Replikation der DNA basiert und einer der grundlegenden Methoden der heutigen Molekularbiologie bildet. Für die Entwicklung wurden beide Forscher mit dem Nobelpreis für Chemie (1993) ausgezeichnet.
Autor: SV
Literatur
Messelson M & Stahl FW (1958) The replication of DNA in Escherichia coli. Proc Natl Acad Sci U S A, vol. 44, pp. 671-682.